Southern route for the East Eurasian Core validated by archaeologic, viral/bacteria, and Ancient DNA data
A short summary on evidence for the southern dispersal route of the EEC, in contrast to the Northern IUP route.
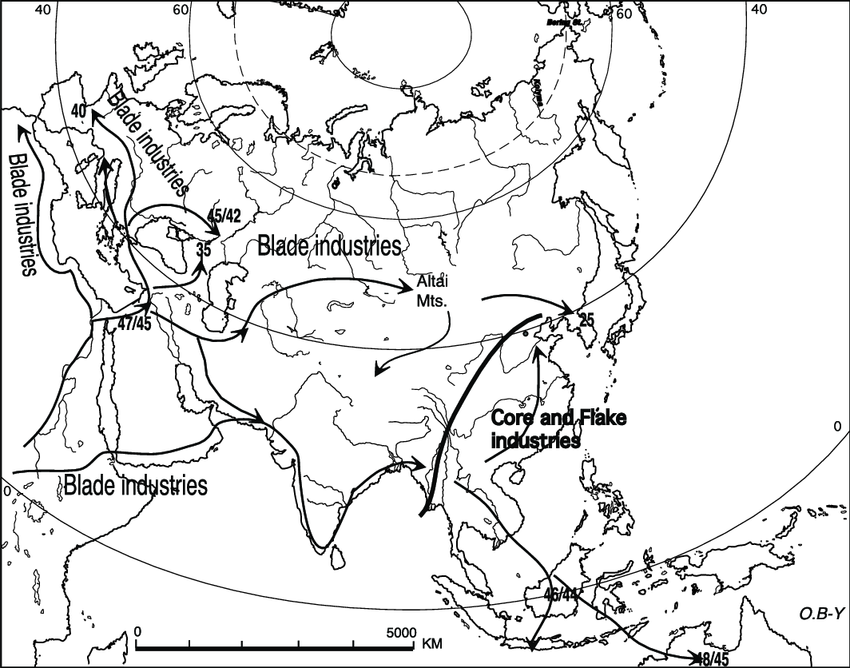
The Northern IUP sites can be associated with a specific microblade material culture, streching from Eastern Europe via Central Asia/Siberia to the Baikal region.
In contrast, Eastern Asia, Southeast Asia, Australasia and South Asia are characterized by a different set of core & flake material culture, representing the southern route. A particular link between East Asian, Tibetan and Hoabinhian core & flake types has been noted.
Those two types are associated with the dispersal of Ancient East Eurasians (c. 48kya) from the eastern Persian plateau. The northern type spreaded with an early diverged lineage or lineages, represented by the genomes of the Ust'Ishim man, the Oase specimens, the Bacho Kiro specimen, as well as sites in the Baikal region and northeastern Uzbekistan. This lineage(s) appearently went largely extinct without substantial contributing to modern populations; there is evidence for geneflow from such northern IUP (Bacho Kiro-like) ancestry into the 39,000 year old Tianyuan specimen from Northern China, as well as geneflow from a similar source into the 35,000 year old GoyetQ116-1 specimen in Europe, further evidenced by the "unusual" affinity observed for Tianyuan, GoyetQ116-1 and BK_IUP. The microblade material culture surrived the extinction of its initial lineage, and was preserved among later Siberian groups, also spreading into the Japanese archipelago and the Americas via ANE and APS.
In contrast, the southern EEC lineages (associated with IUP core & flakes) form the main ancestral source for modern Asia-Pacific populations, having rapidly expanded and diverged along the Southern route into South-Southeast Asian, subsequently populating Oceania and Eastern Asia.
The oldest EEC specimen to date is the Tianyuan man from Northern China, found within a core & flake site. Genetically, the Tianyuan specimen can be modeled as primarily Onge-like + northern IUP (Bacho Kiro-like). In tandem, ancient and modern East Asians form either a trifurication between the Proto-Tianyuan and Onge lineages, or are an early admixture between a primarily Onge-like source + a Tianyuan-like source. One best-fit qpAdm model gave 79% Onge and 21% Tianyuan for AEA. ANEA and Jomon displayed minor additional Tianyuan input, while Longlin/Guangxi had AEA+minor extra Onge/Hoabinhian.
The distribution of northern IUP microblade sites and southern IUP core & flakes, and inferred migration routes:

Core and Flakes were widely distributed in the East and Southeast Asia region, Australasia and much of South Asia. IUP microblades were primarily found in Central Asia/Siberia, with some sites in South Asia and on the Tibetan plateau.
This dual Eastern Eurasian human disperal has been noted before (for example Bar-Yosef & Belfer-Cohen 2013), yet Ancient DNA analyses were missing to determine the respective affilations:
Now, over the last years, we gained more insights via several genetic studies and increased sampling numbers. – The present genetic data revealed that the population of northern IUP sites, such as the Ust'Ishim specimen or the Bacho Kiro specimen, were all basal to modern East Eurasians (AASI, East Asian & Onge, Australasians, and UP Tibetans etc.), but not ancestral to them, representing a deep divergence (48–46kya) from them, shortly after having diverged from West Eurasians at c. 50kya (represented by the UP Kostenki14 and Sunghir remains).
Modern East Eurasian populations are all closer to each other than to those northern IUP specimens (hence referred to as "East Eurasian Core"). The only exception is the Tianyuan specimen, who, although belonging to the southern route wave (EEC), displays increased affinity to the Bacho Kiro/Oase specimens, as well as the GoyetQ116-1 specimen (who is of largely West Eurasian ancestry ~77%). The reason for that is northern IUP (Bacho Kiro-like) geneflow into those two specimens at the respective other end of Eurasia.
Combining archaeogenetic data and archaeologic evidence on the spread of different types of material culture during the IUP/UP, we get a clearer picture on the population history of Eurasia and the phylogenetic structure of early and modern East Eurasians:

For this also compare Vallini et al. 2022 and 2024~
Beyond that, viral/bacteria data support a single southern origin for the ancestors of modern East Asians and other living East Eurasians, aligning well with the archaeogenetic and archaeologic data: Aoki et al. 2023 "Infectious diseases may have arrested the southward advance of microblades in Upper Palaeolithic East Asia".
A single major migration of modern humans into the continents of Asia and Sahul was strongly supported by earlier studies using mitochondrial DNA, the non-recombining portion of Y chromosomes, and autosomal SNP data [42–45]. Ancestral Ancient South Indians with no West Eurasian relatedness, East Asians, Onge (Andamanese hunter–gatherers) and Papuans all derive in a short evolutionary time from the eastward dispersal of an out-of-Africa population [46,47]. The HUGO (Human Genome Organization) Pan-Asian SNP consortium [44] investigated haplotype diversity within present-day Asian populations and found a strong correlation with latitude, with diversity decreasing from south to north. The correlation continues to hold when only mainland Southeast Asian and East Asian populations are considered, and is perhaps attributable to a serial founder effect [50]. These observations are consistent with the view that soon after the single eastward migration of modern humans, East Asians diverged in southern East Asia and dispersed northward across the continent.
...
What stopped the southward expansion of the northern foragers with microblades? We propose and investigate the hypothesis that mortality (and/or infertility) due to infectious diseases prevalent in the south was the cause. After reviewing the literature on the genetic aspects of the peopling of East Asia by modern humans and the temporal–spatial distributions of several infectious diseases during the Palaeolithic, we formulate simple mathematical models that make explicit the structure of our hypothesis. Specifically, we use an approach based on wave-of-advance/reaction–diffusion models [22–26], which permits us to numerically obtain the conditions for the existence of a static disease barrier.
The models suggest that, if each southern forager were to have infected and caused the death of one in a thousand northern foragers per year, then the disease barrier could have been effective in arresting the southward spread of microblades.
In this regard, we have a solidified model on the peopling of the Asia-Pacific route, explaining the distribution of different types of material culture, and unresolved genetic affinities between ancient specimens and modern populations.
The northern IUP microblade material culture spreaded mainly via a northern route into Central Asia, Siberia and eastern Europe, being associated with deep/basal East Eurasian lineage(s) such as represented by the Ust'Ishim specimen or the Bacho Kiro/Oase specimens.
The Southern IUP/EEC wave with Core and Flake material culture, spreaded via the southern route into South Asia, Southeast Asia, East Asia and Oceania. This lineage is ancestral to modern-day East Eurasian populations (AASI, ESEA, Australasians,...) in the Asia-Pacific region.
For a mode detailed take-on on the phylogenetic structure of Ancient East Eurasians, see my first blog post.
%20(1).jpg)

This comment has been removed by the author.
ReplyDelete